Immune Responses to Infectious Diseases Overview

This content delves into immune responses to infectious diseases, covering topics such as barriers and vectors, the link between location and immune effector mechanisms, and methods used by pathogens to evade host defenses. It emphasizes the importance of understanding how pathogens breach barriers, interact with host cells, and trigger immune responses.

Download Presentation

Please find below an Image/Link to download the presentation.
The content on the website is provided AS IS for your information and personal use only. It may not be sold, licensed, or shared on other websites without obtaining consent from the author.If you encounter any issues during the download, it is possible that the publisher has removed the file from their server.
You are allowed to download the files provided on this website for personal or commercial use, subject to the condition that they are used lawfully. All files are the property of their respective owners.
The content on the website is provided AS IS for your information and personal use only. It may not be sold, licensed, or shared on other websites without obtaining consent from the author.
E N D
Presentation Transcript
Immune Responses to Infectious Immune Responses to Infectious Diseases Diseases Dr. Refif S. Al-Shawk Lec 2 2021-2022
Learning Objectives After reading this chapter, you should be able to: 1. Apply previous immunology knowledge when presented with a new pathogen, including location of infection and specific pathogen characteristics, to identify specific innate immune response elements that would be most effective in the initial recognition of the infection, and adaptive response elements (e.g., antibodies versus Tc cells) most appropriate for detection and elimination. 2. Describe and categorize the various methods used by different infectious organisms to evade host immune defense, and specifically explain how each subverts particular elements of host immunity.
The Importance of Barriers and Vectors in Infectious Disease The Importance of Barriers and Vectors in Infectious Disease In order for a pathogen to establish an infection in a susceptible host It must breach physical and chemical barriers Epithelial surfaces of the skin lining of the gut. Is it easy or difficult ?? The difficulty of penetrating these surfaces ensures that most pathogens never gain productive entry into the host. Why? gastric enzymes by specialized epithelial cells lowers the pH of the stomach and upper gastrointestinal tract specialized cells in the gut produce antibacterial peptides. normal commensal flora present at mucosal surfaces (the gastrointestinal, urogenital, and respiratory tracts) can competitively inhibit the binding of pathogens to host cells. Sometimes infectious agents get help from other organisms to circumvent host barriers. In these cases a third party, called a vector, helps to carry the infection from one organism to another. intermediate hosts, can transmit a pathogen between one infected human and another, or from an infected animal to a human. (blood-sucking arthropods (e.g., ticks, fleas, flies, or mosquitoes), breach natural barriers like the skin with their bite and introduce the pathogens they carry directly into a susceptible host.
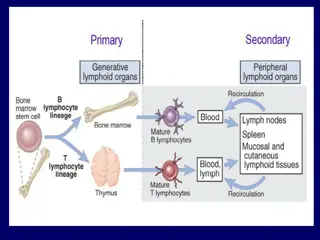
The Link between Location and Immune The Link between Location and Immune Effector Mechanism Effector Mechanism There are many different places that infectious agents can be IN or ON the body. Skin or mucosa or they can penetrate epithelial layers, breaching skin, urogenital, or gut linings. Once they have crossed these barriers, infectious agents may remain local or spread from the site of entry. Some infectious agents can be found in the interstitial fluid that bathes our tissues, where they should then be carried to draining lymph nodes. Infections that actually enter the bloodstream (called sepsis) are rare. When this happens, the infection can spread quickly throughout the body, and the resulting immune response can do more damage than the pathogen itself (e.g., septic shock). Finally, some pathogens spend all or part of their lives inside host cells, occupying either membrane-enclosed (vesicular) or cytosolic and nuclear spaces. Importantly, the entry site and ultimate location of an infectious agent IN or ON the body will determine which immune tools are available and best suited for pathogen detection and elimination.
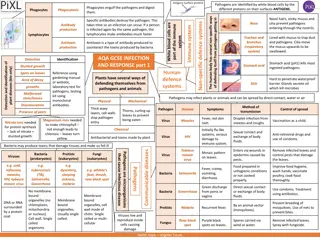
Most infectious organisms enter through: mucosal surfaces (M), like the respiratory, gastrointestinal, and urogenital tracts. Alternatively, they may breach skin barriers through a wound or via an insect bite. Once they have crossed this barrier they exist in the sterile microenvironment of the body, where they can remain local, or spread through the blood and lymphatic circulation. Examples of all four types of pathogen can be found in the extracellular fluid (E), while intracellular parasites and bacteria may be engulfed by or enter host cells and remain vesicular (V). Viruses fuse with the plasma membrane and enter the cytosol (C), from where they may insert their genetic material into host chromosomes in the nucleus. Each of these locations allows for access to different detection necessitate different effectors for elimination. molecules and may
Mucosal or Barrier Infections Are Typically Mucosal or Barrier Infections Are Typically Controlled by T H 2 Controlled by T H 2- -Type Responses Type Responses The majority of, but not all, infectious agents enter through mucosal routes: the mouth, nose, eyes, or urogenital Dangerous infectious agents need to be limited or eradicated before they can induce damage or penetrate the epithelial cell lining of the body, while still maintaining tolerance and even collaboration with commensals. The immune effectors available here include soluble antimicrobial proteins secreted by epithelial cells, innate lymphoid cells, and a dispersed set of lymph node like structures, collectively referred to as mucosal-associated lymphoid tissue (MALT).
Mucosal or Barrier Infections Are Typically Mucosal or Barrier Infections Are Typically Controlled by T H 2 Controlled by T H 2- -Type Responses Type Responses Immunity against many of the most common metazoan parasites (helminths, or worms) correlates with high antigen-specific IgE:IgG ratios. For this reason, cells expressing the high-affinity IgE receptor (e.g., mast cells, basophils, and eosinophils) are also important players in this type-2 response. In some cases, activation of these effector responses leads to expulsion of helminths from body cavities or surfaces. Dimeric IgA (dIgA), serves an important role in neutralizing potential pathogens and maintaining barrier integrity. Binding of IgA to infectious agents can block attachment to epithelial cells (neutralization) and thus aid in elimination of the infectious organism passively, without induction of inflammation [chronic inflammation of the gut correlates with dysbiosis, disruption of barrier integrity, systemic immune imbalance, and inflammatory disease. Thus quiet exit of surface pathogens via noninflammatory means can actually prevent further disease]
Extracellular Pathogens Must Be Recognized Extracellular Pathogens Must Be Recognized and Attacked Using Extracellular Tools and Attacked Using Extracellular Tools When infectious organisms do breach epithelial barriers: Enter the fluid-filled extracellular spaces of the body, namely interstitial fluid or the bloodstream Extracellular infections can remain local or spread through the body, via either the circulatory or lymphatic system. The immune mediators located, include pattern recognition receptors (PRRs) on phagocytic cells, complement, antimicrobial compounds, cytokines (which activate immune cells), and antibody (especially IgG, mIgA, and IgM). Often, T H17, and/or TH 2 cells are the T cell drivers behind the adaptive elements of the response against extracellular infections. All classes of pathogen that breach epithelial barriers will be found in the extracellular spaces (even viruses) for at least some of their lifetime in the host. Extracellular immune effectors play a role in recognition, neutralization, and eradication whenever the pathogen is present
Mechanisms That Recognize Infected Host Cells Are Mechanisms That Recognize Infected Host Cells Are Required to Combat Intracellular Infections Required to Combat Intracellular Infections Infectious organisms that reside inside host cells are the most difficult for the immune response to find and to destroy. bacteria and intracellular parasites and all viruses, which are obligate, intracellular pathogens. Antibodies do not recognize a pathogen hidden inside a cell, a space to which they do not have access, although they can effectively neutralize this infectious agent before it binds to host cells. Most bacteria and parasites that are intracellular spend at least some time within intracellular vesicles, where they might trigger endosomal PRRs, such as certain TLRs (Toll-like receptors). Eradication of infectious organisms that occupy endosomal spaces inside host cells requires a strong TH1 pathway response. This classic delayed-type hypersensitivity (DTH) response relies on cytokines secreted by TH1 cells (e.g., IFN- ) to activate macrophages that can digest these vesicle-bound agents and overcome common pathogen-induced immune evasion of phagosomelysosome fusion. Mycobacterium species, including the agent that causes tuberculosis, are an example
Mechanisms That Recognize Infected Host Cells Are Required to Mechanisms That Recognize Infected Host Cells Are Required to Combat Intracellular Infections Combat Intracellular Infections All viruses and some parasitic agents eventually enter the cytosol of their host cell, where they will spend much of their time Cytosolic PRRs such as the NOD-like receptors (NLRs) and RIG-I-like receptors (RLRs) recognize these agents. This leads to cytokine secretion, sometimes associated with inflammasome activation, and ultimately the induction of cytotoxic cells capable of killing infected host cells; namely, NK cells and CTLs. HOW? NK cells can kill infected host cells via antibody-dependent cell-mediated cytotoxicity or by detecting cell-surface changes that are warning signs of infection (e.g., decreased MHC class I expression or changes in the expression of killer inhibitory receptors). CTLs, activated by cross-presentation of antigen by TH1-licensed DCs, are the primary adaptive mediators of infected target cell killing. In some instances, cytotoxic TH1 cells may also kill infected target cells via Fas Fas ligand binding.
Viral Infections Viral Infections Require host resources for their replication. Passage across the mucosa of the respiratory, urogenital, or gastrointestinal tract accounts for most instances of viral transmission Viruses may also gain entrance through broken skin, such as during an insect bite or puncture wound.
The Antiviral Innate Response Provides Key The Antiviral Innate Response Provides Key Instructions for the Later Adaptive Response Instructions for the Later Adaptive Response A number of specific innate immune effector mechanisms, together with nonspecific defense mechanisms, can prevent or eliminate many viral infections these include: Complement activation via innate pathways and antimicrobial peptides Recognition of pathogen associated molecular patterns (PAMPs) by PRRs expressed by phagocytic cells induces the expression of type I interferons (IFN- and IFN- ) assembly of intracellular inflammasome complexes The binding of type I interferon to NK cells induces lytic activity, making them very effective in killing virally infected cells This activity is enhanced by IL- 12, a cytokine that is produced by dendritic cells very early in the response to viral infection.
Many Viruses Are Neutralized by Antibodies Many Viruses Are Neutralized by Antibodies Neutralizing antibodies, especially those at the sites of infection, as well as circulating antibodies that foster opsonization, complement activation, and phagocytosis, protect the host by blocking or eliminating virus in the extracellular spaces, although they cannot eliminate virally infected cells. Secretory IgA Blocks binding of virus to host cells thus preventing infection or reinfection IgG, IgM, and IgA antibody Block fusion of viral envelope with host cell s plasma membrane IgG and IgM antibody Enhance phagocytosis of viral particles (opsonization) IgM antibody Agglutinates viral particles Complement activated by IgG or IgM antibody Mediates opsonization by C3b and lysis of enveloped viral particles by membrane-attack complex
Cell Cell- -Mediated Immunity Is Important for Viral Mediated Immunity Is Important for Viral Control and Clearance Control and Clearance In order to eliminate an established infection, where host cells harbor intracellular virus. Virus specific CD8 T cells must be activated to kill infected cells, which requires the assistance of helper T cells (often TH1 type) that recognize the same pathogen, providing cytokines and APC licensing for cross-presentation. Cytokines, including IL-2, IFN- , and TNF- , which defend against viruses either directly or indirectly: IFN- acts directly by inducing an antiviral state in nearby cells. IL-2 acts indirectly by helping to activate CTL precursors, generating an effector population of cytotoxic cells. Both IL-2 and IFN- activate NK cells, which play an important role in host defense and lysis of infected cells, especially prior to specific CTL responses.
The Antiviral Innate The Antiviral Innate Response Provides Response Provides Key Instructions for Key Instructions for the Later Adaptive the Later Adaptive Response Response
Viruses Employ Several Strategies to Evade Host Defense Viruses Employ Several Strategies to Evade Host Defense Mechanisms Mechanisms Evasion the action of IFN- / . For instance, hepatitis C virus overcomes the antiviral effect of the interferons by blocking or inhibiting the action of PKR, a protein kinase essential to signal transduction Inhibition of antigen presentation by infected host cells. Herpes simplex virus (HSV) produces a protein that inhibits the human transporter molecule needed for antigen processing (TAP) Inhibition of TAP blocks antigen delivery to MHC class I molecules and effectively shutting down presentation of HSV antigens to CD8+ T cells and CTL recognition of infected cells. Adenovirus and cytomegalovirus (CMV) use distinct molecular mechanisms to reduce the surface expression of MHC class I molecules, again inhibiting antigen presentation to CD8 T cells. A number of viruses escape immune attack by constantly changing their surface antigens. Like influenza virus and HIV. Some viruses, such as Epstein-Barr virus (EBV) and HIV, can cause generalized or specific immunosuppression: In the case of HIV, viral infection of lymphocytes or macrophages destroy the immune cells or alter their function. Cytokine imbalance or pathogen-induced diversion toward less effective immune response pathways. For instance, EBV, the cause of mononucleosis, produces a protein that is homologous to IL-10; like IL- 10, this protein suppresses cytokine production by the TH1 subset, resulting in inhibition of the antiviral inflammatory response.
Bacterial Infections Bacterial Infections Bacteria can enter the body through a number of natural routes (e.g., the respiratory, gastrointestinal, and urogenital tracts) or through normally inaccessible routes opened up by breaks in mucous membranes or skin. Depending on the number of organisms entering and their virulence, different levels of host defense are enlisted. If the inoculum size and the virulence are both low, then localized tissue phagocytes may be able to eliminate the bacteria via nonspecific innate defenses. Larger inocula, organisms with greater virulence, and intracellular bacteria typically require antigen-specific adaptive immune responses. 4 steps: Attachment to host cells Proliferation Invasion of host tissue Toxin-induced damage to host cells
Immune Responses to Extracellular and Intracellular Immune Responses to Extracellular and Intracellular Bacteria Differ Bacteria Differ Immunity to bacterial infections is usually achieved by a combination of humoral and cell-mediated immunity, depending on the type of pathogen. humoral immune response is the main protective response against extracellular bacteria. Antibodies which are secreted by plasma cells in regional lymph nodes or the submucosa of the respiratory and gastrointestinal tracts 1. Antibody neutralizes bacterial toxins. 2. Complement activation on bacterial surfaces leads to complement-mediated lysis of bacteria. 3. Antibody and the complement split product C3b bind to bacteria, serving as opsonins to increase phagocytosis. 4. C3a and C5a, generated by antibody-initiated complement activation, induce local mast- cell degranulation, releasing substances that mediate vasodilation and extravasation of lymphocytes and neutrophils. 5. Other complement products are chemotactic for neutrophils and macrophages.
Immune Responses to Extracellular and Intracellular Bacteria Differ
Intracellular (vesicular) bacteria present a different challenge due to their residency inside host cells. Activation of TLRs in membranes Also activate NK cell mediated killing which in turn provides an early defense against these organisms. Ultimately, to be effective, the cellular response to intracellular bacteria requires TH1 cell mediated immune responses, such as DTH In this response, IFN- secreted by CD4 T cells are crucial for activating macrophages to phagocytose and kill these bacteria more effectively. The most notable example of this is in the Mycobacterium family of intracellular pathogens, where strong TH1 responses have been shown to protect against or clear this infection
Bacteria Can Evade Host Bacteria Can Evade Host Defense Mechanisms at Defense Mechanisms at Several Different Stages Several Different Stages Including attachment to host cells, blocking IgA, inhibiting complement changing antigenic structures inhibiting phagocytosis or phagosome-lysosome fusion and thus avoiding intracellular degradation.
Referrences Referrences : : Kuby immunology 8th edition, 2019 Cellular and molecular immunology 9th edition , 2018 by Abul K. Abbas






















 bacteria present a different")
 bacteria present a different")